Asociacija (ARBA „KONCATENACIJA“ ARBA „NUORODA“)
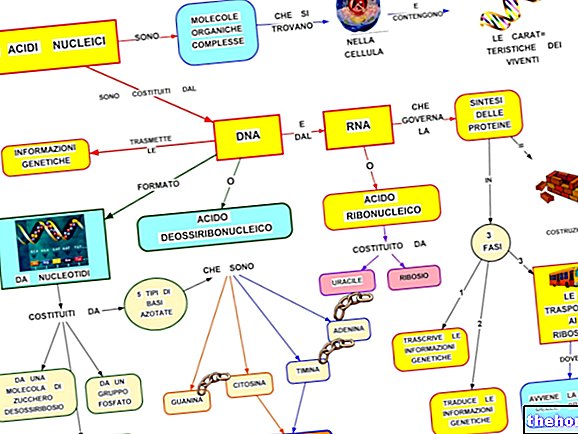
Iki šiol mes gydėme Mendelio dihibridinį (arba polihibridinį) kirtimą darant prielaidą, kad skirtingos alelinės poros iš tikrųjų randamos skirtingose homologinėse chromosomų porose. Tačiau chromosomų porų skaičius, nors ir skiriasi skirtingoms rūšims, kinta ribotai (nedaug rūšių) pasiekia apie šimtą chromosomų), o genų skaičių galima suskaičiuoti dešimtimis tūkstančių.
Kad Mendelio savo eksperimentams pasirinkti simboliai atsiskyrė visi nepriklausomai (todėl nesupainiojami skaičiavimai, susiję su fenotipų pasiskirstymu daugiakrybio kryžiaus F2), buvo laiminga galimybė. Jei gretimuose lokusuose būtų rastos dvi alelių poros, įstatymas būtų vadinama asociacijos įstatymu.
Žinant, kad labai daug simbolių turi savo lokusą vienoje chromosomoje ir kad jie yra chromosomų poros, kurios mejozėje atsiskiria nepriklausomai, galima suprasti, kaip dažnai atsitinka, kad dvi poros simbolių, jei jie buvo susieti su tėvų chromosoma taip pat ir lytinėse ląstelėse, taigi ir organizme, į kurį jis atneš savo genetinę medžiagą.
Taigi matome, kad „asociacija yra išimtis“, toli gražu ne reta, nepriklausomumui, įtvirtintam trečiajame Mendelio įstatyme.
KEITIMAS ARBA „PERKLAVIMAS“ IR REKOMBINACIJA
Kalbėdami apie mejozę, mes nurodėme, kad yra du skirtingi genetinės medžiagos maišymo momentai: vienas yra chromosomų atskyrimas lytinėse ląstelėse, tą pastebėjo Mendelis.
Kitas momentas, kuris iš tikrųjų buvo ankstesnis, yra tas, kai kiekvienos homologinių chromosomų poros keturios chromatidės tarpusavyje keičiasi identiškais bruožais. Po šio keitimosi du veiksniai, susieti su ta pačia chromosoma, bus nepriklausomi nuo lytinių ląstelių. Tikimybė kad mainai vyksta, yra proporcingas, pirma apytikslė, chromosomos ilgiui, o ilgesnėse chromosomose gali būti net daugiau nei, mainai.
Šį reiškinį galima aptikti citologiškai, stebint pakankamą skaičių mejozių mikroskopu.
Pakartotinių atitikties rodiklis yra greitis, kuriuo bet kurie du simboliai, susieti tėvų kartoje, F2 skiriasi.
Jei abu lokusai yra absoliučiai besiribojantys, tikimybė, kad chiasmas juos atskirs, bus praktiškai lygi nuliui. Rekombinacijos greitis bus: n ° rekombinantų. Jei du lokusai yra dviejose skirtingose chromosomose, rekombinacijos dažnis bus 0,5 (lygi tikimybė, kad du simboliai, sujungti P kartoje, atsitiktinai atsidurs kartu F2). Todėl rekombinacijos dažnis gali svyruoti nuo 0,0 iki 0,5. Mažiems atstumams chromosomoje atstumas ir rekombinacijos greitis yra tiesiogiai proporcingi. Ilgesniems atstumams yra galimybė, kad tarp dviejų lokusų vyksta du mainai. Dabar bus aišku, kad du veiksniai, atskirti dviem mainais, vėl yra susiję. šiuo metu akivaizdu, kad prarandamas proporcingumas tarp lokusų atstumo ir rekombinacijos tikimybės.
Lokusai, rasti toje pačioje chromosomoje, sudaro „asociacijos grupes“. Labai tolimi lokusai gali turėti tokią tikimybę atskirti mainus, kad jie elgiasi kaip nepriklausomi, tačiau kiekvienas iš jų bus susietas su mažesniu rekombinacijos greičiu su tarpiniais lokusais.
Kai žinomos daugelio asociacijos grupės genų porų rekombinacijos normos, galima pradėti kurti „genetinius žemėlapius“. Turint omenyje, kad atstumas tarp dviejų genų (a ir b) išreiškiamas rekombinacijos greičiu ir kad a atstumas nuo trečiojo geno c gali būti suma arba skirtumas nuo jo atstumo nuo b, tai įmanoma rekonstruoti abipusių atstumų žemėlapį, kuris bus tos asociacijos grupės, ty tos chromosomos, genetinis žemėlapis.
Dabar turime apskritai apsvarstyti kai kurias sąvokas, kurios riboja fenotipinį genotipinių simbolių pasireiškimą.
Pirmiausia kalbėsime apie skvarbumo ir išraiškingumo sąvokas, o tada ypatingą dėmesį skirsime genų veikimo reguliavimo reiškiniams.
PENETRANSAS
Geno įsiskverbimas reiškia jo gebėjimą pasireikšti fenotipuose. Skvarbumas matuojamas statistiškai, skaičiuojant fenotipų, rodančių tą charakterį, dažnį iš 100 genotipų, kuriuose jis yra. Bruožas, kurio skvarbumas yra 0,7, yra bruožas, kuris fenotipiškai pasireiškia 70% jo genotipinio dažnio.
IŠRAIŠKUMAS
Išraiškingumas yra kiekybinis fenotipinio pasireiškimo laipsnio įvertinimas.
GENO VEIKSMŲ REGULIAVIMAS
Ląstelės gamina visus savo fermentus ir baltymus tuo pačiu greičiu ir tuo pačiu metu. Pavyzdžiui, Escherichia coli ląstelės gali būti aprūpintos energijos ir anglies atomais iš laktozės disacharido, nes jos gali suskaidyti jas į gliukozę ir galaktozę dėl beta-galaktozidazės fermento.coli, kuriame gali būti laktozės, yra maždaug 3 000 beta-galaktozidazės molekulių, atitinkančių 3% tos ląstelės baltymų; nesant laktozės, vienoje bakterijos ląstelėje bus tik viena beta-galaktozidazės molekulė. mRNR molekulės, kai jas galima naudoti. Mutantinės E. coli padermės, kuriose gausu fermento, yra žinomos net tada, kai nėra laktozės: šie mutantai yra nepalankioje padėtyje, palyginti su normaliomis ląstelėmis, nes yra priversti be reikalo sunaudoti energijos ir medžiagų. fermento, kuris liks be substrato. Medžiagos, dėl kurių padidėja fermento kiekis, kaip ir laktozės atveju, bus vadinamos induktoriais, o fermentai - indukuojamais. Kitos medžiagos, net ir būtent šios, sukelia gamybą tam tikrų fermentų. Pavyzdžiui, E. coli, galintis gaminti visas savo amino rūgštis, turinčias anglies ir amonio (NH3), Tam tikros aminorūgšties (pavyzdžiui, histidino) buvimas auginimo terpėje blokuoja visų fermentų, susijusių su pačios aminorūgšties biosinteze, gamybą: apie šiuos fermentus bus pasakyta, kad jie yra slopinami. Bakterijų ląstelėse mRNR molekulės suskaidomos netrukus po jų susidarymo, todėl kontroliuoti mRNR gamybą reiškia kontroliuoti fermentinę sintezę tuo pačiu metu.
OPERONAS
Norėdami paaiškinti, kaip bakterijų ląstelė sugeba kontroliuoti savo fermentų gamybą, Jokūbas ir Monodas suformulavo operono hipotezę; operoną sudaro keli genai, kurie yra funkciškai susiję ir be pertrūkių suderinti išilgai DNR ruožo.Operonas susideda iš trijų skirtingų tipų genų: promotoriaus, kuriame prasideda mRNR susidarymas; operatorius, kur kontroliuojama; vienas ar daugiau struktūrinių genų, koduojančių fermentus ar kitus baltymus. Beta-galaktozidazės sistemoje operonas, be beta-galaktozidazės, taip pat apima du kitus genus, kiti fermentai, dalyvaujantys laktozės metabolizme. Šie genai yra greta vienas kito ir vienas po kito perrašomi išilgai tos pačios DNR spiralės į vieną mRNR molekulę. Taip pagamintos mRNR molekulės veikia labai trumpą laiką, po to jas sunaikina specifiniai fermentai.
Operono veiklą savo ruožtu kontroliuoja kitas genas - reguliatorius, kuris taip pat gali būti nutolęs nuo operono: šis reguliatorius koduoja baltymą, vadinamą represoriumi, kuris, atrodo, jungiasi prie operatoriaus geno DNR. tarp promotoriaus ir struktūrinių genų iš tikrųjų blokuoja mRNR gamybą.
Savo ruožtu represorius yra valdomas, o valdymas atliekamas naudojant „signalinę“ medžiagą. Indukuojamų fermentų atveju ši medžiaga yra "induktorius. Induktorius" jungiasi prie represoriaus molekulės, pakeisdamas jos formą, kad ji nebegalėtų prisitaikyti prie DNR: šiuo atveju, kadangi tarp promotoriaus ir struktūrinių genų nėra represoriaus , represorius gali sudaryti mRNR molekules ir iš jų baltymų molekules. Pasibaigus induktoriaus tiekimui, reguliatorius vėl atgaus kontrolę, o tai sustabdys naujų mRNR, taigi ir naujų baltymų, gamybą. Beta-galaktozidazės sistemoje induktorius yra laktozė arba į ją labai panaši medžiaga. Darinys: jie prisijungs prie represoriaus, jį inaktyvindamas, kad būtų leista fermentų biosintezė. Esant represuojamiems fermentams, medžiaga, kuri veikia kaip „signalas“, veikia kaip šerdies slopintuvas: represorius yra aktyvus tik tada, kai yra derinamas su pagrindiniu. Histidino sistemoje, kurioje dalyvauja keliolika skirtingų fermentų, būtent ši aminorūgštis, kartu su jos tRNR, korepresoriumi histidinu.
ALOSTERINĖ SĄVEIKA
Allosterinė sąveika, apimanti fermento inaktyvavimą keičiant jo formą, suteikia kitokį būdą reguliuoti ląstelės metabolinį aktyvumą. Allosterinė sąveika leidžia tiksliau kontroliuoti nei operono induktoriaus -represoriaus sistema, tačiau nepasiekiamas naudingas rezultatas, kai tam tikros medžiagos biosintezė pašalinama iš pirmojo etapo - mRNR gamybos.
KONTROLĖS SISTEMOS EUCARIUS
Yra keletas faktų, leidžiančių manyti, kad panaši į operoną reguliavimo sistema veikia ir vyrauja tarp augalų ir gyvūnų. Šių organizmų chromosomos labai skiriasi nuo E. coli ir kitų prokariotų. Šių genų kontrolė mitozės mechanizmas yra toks, kad kiekviena tam tikro augalo ar gyvūno ląstelė turi visą informaciją
genetika, esanti apvaisintame kiaušinyje. Todėl dauguma genų bet kurioje specializuotoje ląstelėje išliks neefektyvūs visą ląstelės gyvenimą. Šių ląstelių DNR visada yra susijusi su baltymais. Taigi gali būti, kad genų slopinimas eukariotuose reikalauja būtent šios asociacijos tarp DNR ir baltymų.






-quando-preoccuparsi.jpg)