Ciklinis gyvų būtybių tęstinumas atranda ryšius tarp vienas po kito einančių kartų reprodukcijos reiškiniuose.
Dauginimas atliekamas skirtingais evoliucinės skalės lygmenimis, skirtingomis augalų ir gyvūnų karalystės pasekmėmis, skirtingomis gyvomis rūšimis, naudojant tokius mechanizmus, kurie vien pateisina visą traktatą.
Pirmoji reprodukcijos reiškinių klasifikacija turi atskirti vienaląsčius nuo daugialąsčių organizmų, nes tik pirmajame ląstelių dalijimasis sutampa su dauginimu.
Daugialąsčių reprodukcija gali būti agaminė arba seksualinė (arba gama).
Agaminis dauginimasis, palyginti rečiau, grindžiamas mitozės mechanizmu, todėl rūšies kintamumas yra patikėtas pakartotiniam mutacijų atsiradimui.
Taip pat yra įvairių mechanizmų, tokių kaip strobilizacija, sporuliacija ir kt., Tuo tarpu augaluose randame žemės ūkyje gerai žinomų regeneracinių formų (auginiai, sluoksniavimas ir kt.).
Tačiau labiausiai paplitęs reprodukcinis mechanizmas aukštesnėse formose yra seksualinis, atitinkantis mejozės atsiradimą, lytinių ląstelių susidarymą ir jų susiliejimą zigotoje (apvaisinimas).
Primityviose rūšyse lytinės ląstelės nėra morfologiškai diferencijuotos: šiuo atveju kalbame apie izogametiją. Tačiau visada yra dvi gametų serijos, identifikuojamos simboliais (+) ir (-), o apvaisinimas gali vykti tik iš „priešingo ženklo lytinių ląstelių susidūrimo: todėl egzistuoja biologinis skirtumas, dar neišreikštas morfologiškai.
Tobulėjant evoliuciniam masteliui, atsiranda morfologinė ir funkcinė diferenciacija, kai moterų lytinių ląstelių tipas paprastai aprūpinamas gausia atsargine medžiaga (deutoplazma arba veršelis, kuris užtikrins embriono vystymąsi, kol jis bus nepriklausomas nuo metabolizmo) ir tam tikros rūšies. vyriškos lytinės ląstelės, turinčios judrumą pasiekti moteriškas. Gametos visada yra haploidinės ir yra mejozės rezultatas. Jų susiliejimas sukelia diploidinę zigotą.
Tarp meozės ir zigotos gali praeiti haploidinių ląstelių kartų serija, lygiai taip pat, kaip tarp zigotos ir mejozės, gali praeiti diploidinių ląstelių kartų serija, o skirtingų generacijų ciklų atmainos išreiškiamos kartų kaita.
Diplonte rūšiai (su diploidiniu organizmu) būdinga gametinė mejozė: mejozė tiesiogiai gamina gametas, kurios, susiliejusios, iš karto atkuria diploidinę būseną. Tai yra paplitęs metaziečių atvejis, įskaitant žmogų.
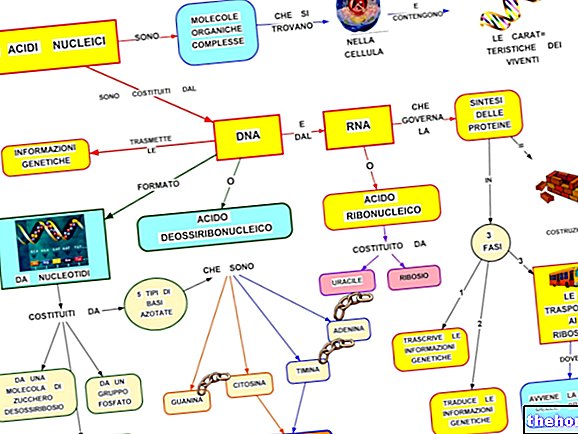
Gametogenezė
Atsižvelgiant į reprodukciją naudojant gametinę mejozę, kaip ji pasireiškia žmogui, pabandykime išsiaiškinti, kaip mejozė dera prie gametogenezės (lytinių ląstelių susidarymo).
Vystantis embrionui, vyriškos ir moteriškos lyties gametogenezei (vadinamai spermatogeneze ir oogeneze), anksti atsiskiria ląstelės, skirtos kūnui formuoti (somatinė linija), ir ląstelės, gaminančios lytines ląsteles (gemalo linija). Pradinės gemalo linijos ląstelės vadinamos protogonais. Skiriant lytines liaukas vyriška ar moteriška prasme, lytinės ląstelės atitinkamai diferencijuojasi į spermatogoniją ir ovogoniją.
Žvelgdami į spermatogenezę, matome, kad spermatogonijoje yra ląstelių kartos, kurios tęsiasi visą gyvenimą. Tik dalis nuolat gaminamos spermatogonijos skiriasi nuo įprasto mitozinio ciklo ir prasideda mejozinis.
Lytinė ląstelė, kurioje prasidės mejozė (reduplikacija, o paskui pirmasis dalijimasis), vadinama pirmos eilės spermatocitu; jo dalijimosi metu susidaro du antros eilės spermatocitai, o su antruoju dalijimusi iš viso atsiranda keturi spermatidai.
Chromosomų rinkinio sumažinimą galime baigti nuo 4 n pirmos eilės spermatocitų (po pakartotinės dubliavimosi kiekvienai homologų porai yra keturios chromatilės) iki 2 antros eilės spermatocitų ir iki spermatidų n, kaip jau matyti mejozė, kurią Taigi darome išvadą. Todėl spermatidės jau yra haploidinės, tačiau jos dar nėra subrendusios lytinės ląstelės. Iš haploidinio tipo ląstelės struktūros funkcinis brendimas (vadinamas spermiohistogeneze) spermatidus paverčia spermatozoidais, tai yra, subrendusiomis lytinėmis ląstelėmis.
Moterų gametogenezėje (arba oogenezėje) yra keletas skirtumų. Visų pirma, ruošiamų lytinių ląstelių skaičius yra daug mažesnis. Manoma, kad žmogaus rūšies patelės lytinėse liaukose yra paruošta apie 5 X 105 ovogonų; iš jų tik apie 400 yra suinteresuoti folikulų brendimu ir vėlesniu nuleidimu, ciklu, kuris paprastai veikia tik vieną folikulą per mėnesį vaisingą maždaug 35 metų laikotarpį.
Skirtingas abiejų lyčių lytinių ląstelių skaičius atitinka jau minėtą funkcijų ir elgesio skirtumą: spermatozoidai yra nedideli, judrūs ir daug, atsižvelgiant į poreikį ieškoti kiaušinio ir mažą tikimybę jį rasti; kiaušiniai yra dideli, inertiški ir nedaug, atsižvelgiant į funkciją, užtikrinančią embrionui rezervinę medžiagą ir apsaugą, kurią jiems suteikia vidinis tręšimas (natūraliai, ypač atliekant išorinį apvaisinimą, kiaušialąstės taip pat turi būti gausesnės).
Poreikis aprūpinti lytines ląsteles atsargine medžiaga atitinka „oogenezėje“ mejozės sustabdymo fazę, kurios metu chromosomos yra iš dalies despiralizuotos. Tada stebime vadinamąsias „plunksnų chromosomas“, kuriose serija Ekstraversijos nustato bruožus, kuriais slopinami genai, atsakingi už deutoplazmos sintezę.
Mažesnis moterų lytinių ląstelių skaičius taip pat atitinka tai, kad iš keturių mejozės pagamintų haploidinių ląstelių tik viena gauna visą rezervinę medžiagą ir tampa lytine liauka, o kitos trys (polocitai arba poliniai kūnai), kuriuose yra tik chromosomų medžiaga, negali sukelti zigotų ir embrionų ir yra skirta regresui.
Tręšimas
Tręšimas, tai yra lytinių ląstelių patino ir vyro susidūrimas, gali būti atliekamas labai įvairiai.Gyvūnų karalystėje mes stebime perėjimą nuo išorinio apvaisinimo (lytinės ląstelės, kurioms gresia bet koks pavojus aplinkai, todėl būtinai labai daug abiejų lyčių) į vidinį apvaisinimą, prie kurio tėvų priežiūra dar labiau siejama su žinduolių metaboliniu ryšiu tarp motinos ir vaisiaus .
Apvaisinimas, įvykus priešingos lyties lytinių ląstelių susidūrimui, turi įvykti garantuojant dvi sąlygas: specifiškumą ir unikalumą. Tai yra, turi būti užtikrinta, kad spermatozoidas būtų tos pačios rūšies kaip ir kiaušinis ir kad, patekus pirmajam, kiti nepatektų.
Specifiškumą užtikrina biocheminės akrosomos ir kiaušialąstės paviršiaus savybės. Tiesą sakant, kalbama apie „tręšiamųjų“ ir „antifertilizinų“ reakcijas, kurių specifiškumas yra panašus į fermentų ir substrato susidūrimo specifiškumą.
Apvaisinimo unikalumą garantuoja kiaušialąstės paviršiaus struktūros pakeitimas („žievės reakcija“), prasidedanti pirmosios specifinės trąšų / antifertilizino reakcijos metu; po šios reakcijos pasikeičia kiaušialąstės membrana, todėl kiti ją pasiekę spermatozoidai nebegali pradėti specifinės apvaisinimo reakcijos.
Po apvaisinimo spermatozoidų uodega lieka išorinė nuo kiaušialąstės, o į ją prasiskverbia chromosomų medžiaga. Ši, vadinama „vyriškuoju pronukleumi“, prisijungia prie kiaušialąstės „moteriškojo pronukleus“ ir taip sudaro diploidinį zigotos branduolį.












.jpg)




-cause-sintomi-e-cura.jpg)