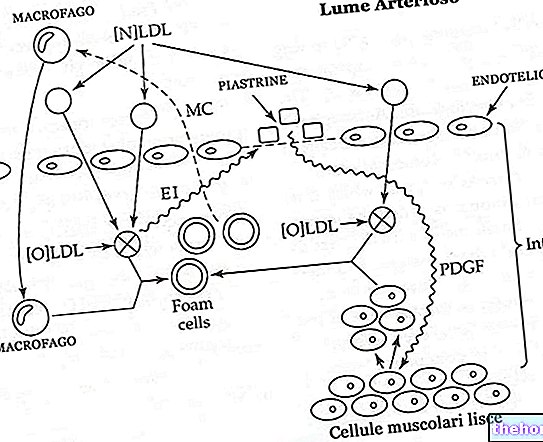
" pirma dalis
Fermentas, katalizuojantis ketvirtą krebso ciklo etapą, yra α-keto glutarato dehidrogenazė; šis fermentas yra fermentų kompleksas, labai panašus į piruvato dehidrogenazę. Abu yra sudaryti iš 48–60 baltymų, kuriuose atpažįstami trys skirtingi fermentiniai aktyvumai, taip pat turi tuos pačius fermentinius kofaktorius; yra labai panašūs fermentai, nes veikia panašius substratus: ir piruvatą, ir l "α-keto glutaratas, yra α- keto rūgštys. Dviejų fermentinių kompleksų veikimo mechanizmas yra tas pats.
Tiamino pirofosfato išpuolis prieš karbonilą (C = O).α-ketoglutaratas, sukelia jo dekarboksilinimą ir susidaro karboksihidroksi propilo darinys. Vėliau perkeliant į lipoamidą, vyksta vidinis redokso procesas, iš kurio gaunamas lipoamido karboksi darinys arba sukcinil-lipoamidas.
Tada sukcinilo lipoamidas reaguoja su kofermentu A ir susidaro sukcinilkoenzimas A (kuris tęsiasi krebso cikle) ir redukuotas lipoamidas, kurį FAD reoksiduoja: susidaręs FADH2 yra reoksiduojamas NAD + ir NADH. Todėl šiame etape įvyko antrasis anglies pašalinimas iš anglies karkaso anglies dioksido pavidalu.
Acilo grupė, susieta su kofermentu A, yra aktyvuota forma, tai yra, ji turi daug energijos: todėl galima išnaudoti sukcinilo kofermento A energiją.

Penktame krebso ciklo etape sukcinilkoenzimas A yra veikiamas sukcinilo tiokinazė; dėl jo veikimo būdo iškeltos dvi hipotezės: aprašysime tik vieną iš dviejų, nes ji yra labiausiai akredituota. Remiantis šia hipoteze, sukcinilo koenzimą A užpuola fermento histidino (Hys) azotas: kofermentas A išsiskiria ir iš histidino gautas adduktas susidaro kaip tarpinė medžiaga, tai yra sukcinilo fermentas (arba sukcinil-Hys) ); šią tarpinę medžiagą veikia ortofosfatas, dėl kurio išsiskiria sukcinatas ir susidaro fosfoenzimas. Fosfoenzimas, užpultas guanozino difosfato (BVP), gamina guaznozino trifosfatą (GTP) ir fermentas išsiskiria. Energetiniu požiūriu GTP = ATP: ryšys, teikiantis energiją, yra vienodas abiejose rūšyse (tai yra anhidridinis ryšys tarp fosforilo Β ir fosforilo γ). Kai kuriais atvejais GTP naudojamas kaip medžiaga, turinti daug energijos, tačiau paprastai GTP veikiant fermentui virsta ATP nukleozido difosfokinazės (NDPK); yra fermentas, randamas ląstelėse ir katalizuoja šią reakciją:
N1TP + N2DP → N1DP + N2TP
Bendras NiTP ® nukleozidų trifosfatas
Bendras NiDP ® nukleozidų difosfatas
Tai grįžtama reakcija; mūsų atveju tai atsitinka:
GTP + ADP → BVP + ATP
todėl jis gali judėti į dešinę arba į kairę net ir esant nedideliems reagentų koncentracijos pokyčiams.
Jei krebso ciklas vyksta tokiu greičiu, kad ATP gamyba viršija energijos poreikį, ADP yra mažai, o ATP yra daug: nukleozidinės difosfokinazės katalizuojama reakcija yra nukreipta į kairę (GTP kaupiasi, jei nukleozidinė difosfokinazė neturi pakankamai substrato, t. y. ADP). Todėl GTP yra energijos prieinamumo signalas, todėl sulėtina krebso ciklą.
Šeštasis krebso ciklo etapas lemia fumarato susidarymą sukcinato dehidrogenazė; šis fermentas sukelia stereospecifinę reakciją, nes nesočiųjų (tai yra alkenas) trans visada susidaro, ty fumaratas (o cis izomeras yra maleatas). Sukcinato dehidrogenazė randama vidinėje mitochondrijų membranoje, o visi kiti krebso ciklo fermentai yra išsibarstę po mitochondriją.
Sukcinato dehidrogenazės kofaktorius yra FAD; jį slopina oksaloacetatas (grįžtamojo ryšio slopinimas), o sukcinatas ir fumaratas yra teigiamas moduliatorius (aktyvatorius). jo aktyvatorius. Pabandykime suprasti, kodėl, pereidami į paskutinį krebso ciklo etapą. Paskutinis etapas krebso ciklas reikalauja energijos, todėl vienintelė galimybė iš paciento gauti oksaloacetato yra ta, kad paciento koncentracija yra labai didelė: malatas yra vienas iš didžiausių metabolitų, turinčių didžiausią koncentraciją ląstelėse. Reakcijai, kuri paverčia malatą į oksaloacetatą, taip pat faktas, kad veikiant citrato sintazei oksalooacetato koncentracija yra maža. Reakcija, kurią katalizuoja sukcinato dehidrogenazė, yra savaime besimaitinanti reakcija, ir tai yra vienintelis būdas paversti malatą į oksaloacetatą.
Mitochondrijų malato koncentracija turi būti suderinama su citoplazminio malato koncentracija: tik tada, kai mitochondrijų malato koncentracija yra tokia didelė, kad garantuojama, kad malatas virsta oksaloacetatu (krebso ciklo metu), tada malatas gali būti naudojamas ir kitame būdai (kurie yra citoplazminiai): citoplazmoje malatas gali būti paverstas oksaloacetatu, iš kurio aspartatą galima gauti veikiant GOT (tai yra transaminazė) arba gliukozei gliukoneogenezės būdu.
Grįžtame prie septintojo krebso ciklo etapo, kurį katalizuoja fermentas fumarasi: vanduo pridedamas stereospecifiniu būdu, kad būtų pagamintas L-malatas.
Paskutiniame Krebso ciklo etape, apie kurį jau kalbėjome, veiksmas malato dehidrogenazė. Šis fermentas kataliziniam veikimui naudoja NAD + molekulę.
Taigi baigėme įvairių krebso ciklo etapų aprašymą.
Krebso ciklas yra visiškai grįžtamas.
Norint padidinti krebso ciklo greitį, galima padidinti tame cikle esančių metabolitų koncentraciją; viena iš strategijų, kaip pagreitinti krebso ciklą, yra dalis piruvato, patenkančio į mitochondrijas, paversti oksaloacetatu (veikiant piruvato karboksilazei), o ne visa tai paversti acetilo kofermentu A: taip padidėja oksaloacetato koncentracija. yra krebso ciklo metabolitas, todėl padidina viso ciklo greitį.
Krebso ciklo metu trys NAD + paverčiami į tris NADH, o vienas FADH - į FADH2 ir, be to, gaunamas GTP: nukreipiant redukcinę galią, gautą iš krebso ciklo, toliau gaminamas ATP; Kvėpavimo grandinėje redukuojanti galia iš NADH ir FADH2 perkeliama į deguonį: šis perkėlimas atsiranda dėl daugybės fermentų, esančių ant mitochondrijų membranos, kurios, savo veikimu, sukelia ATP gamybą.
Kvėpavimo grandinės procesai yra egzergoniniai procesai, o išlaisvinta energija naudojama ATP gamybai; ląstelės tikslas yra išnaudoti eksergoninius procesus, kad vyktų ATP sintezė. Kiekvienai į kvėpavimo grandinę patekusiai NADH molekulei gaunama 2,5 ATP molekulės, o kiekvienai FADH2 - 1,5 ATP molekulės; ši įvairovė atsiranda dėl to, kad FADH2 patenka į kvėpavimo grandinę žemesniu lygiu nei NADH.
Sumažinus aerobinio metabolizmo galią, gaunama 30-32 ATP (219-233 kcal / mol), kurių išeiga yra apie 33% (anaerobinio metabolizmo išeiga yra apie 2%).